Biological Control of Diaprepes abbreviatus by Parasitoids
Jorge E. Peña and D. M. Amalin
University of Florida, IFAS
Tropical Research and Education Center
Homestead, FL 33031
The West Indian sugarcane rootstalk borer weevil, Diaprepes abbreviatus (L.) has been an important pest of several crops (i.e., citrus, sugarcane, vegetables, ornamentals) in Florida for over 30 years. In Florida, annual losses and cost of control in citrus currently exceeds 72 million dollars, while losses in ornamentals and vegetables were close to 2 million dollars during 1997 through 1999. Different pest management tactics are being developed to manage Diaprepes abbreviatus and they include autacoidal and chemical methods, plant resistance and use of biopesticides. Biological control by entomophagous insects is viewed as well suited for management of Diaprepes abbreviatus, because of its minimal physical input, cost-effectiveness and because it offers the potential for long-lasting control of the target pest. However, until today, very little effort has been made to include the use of entomophagous insects in the whole management program for Diaprepes abbreviatus. Entomophagous insects are divided into two general classes the parasitoids and the predators. The term parasitoid, when applied to an entomophagous insect, refers to one that, in its larval stage, develops either internally or externally upon a single host individual, and the host eventually dies as a result of the attack. Parasitoids as adults are, with few exceptions, free-living, and their food sources are usually distinct from those of the larvae (DeBach, 1974).
Biological control of Diaprepes species by parasitoids and predators already present in Puerto Rico, Guadeloupe and Florida has been observed (Table 1) (Whitcomb et al., 1982; Beavers et al., 1980; Peña et al., unpublished data; Etienne et al., 1990). On the other hand, some conflicting reports exist in the literature about adult and larval parasitoids of this pest (Thompson, 1943). For instance, there are reports of the existence of a larval parasitoid of Diaprepes in Brazil, while there is no evidence that Diaprepes is distributed in that area. Thus, there has been short-lived research on parasitoids as control agents of Diaprepes abbreviatus. It is quite important to re-direct efforts to the development of a management system which take all possible advantage of naturally occurring biological control and that considers the possibility of intensive studies of natural enemies in the area of origin of Diaprepes abbreviatus.
Aspects to Consider for a Successful Program on Biological Control of Diaprepes abbreviatus
Life History of Diaprepes abbreviatus
Insects that spend part of their lives in the soil, such as Diaprepes abbreviatus, present a challenge as biological control targets. The life stages that are observed on leaves (i.e., eggs and adults), are considered more exposed to the action of parasitoids than those that are found in the soil (i.e., larvae and pupae). This point of view, however, could be influenced by the fact that eggs and adults are more easily seen by researchers and growers than larvae and pupae of the Diaprepes weevil in the soil. Consequently, more efforts have been directed to collect parasitoids of Diaprepes eggs than from pupae, larvae or even adults.
Egg Parasitoids
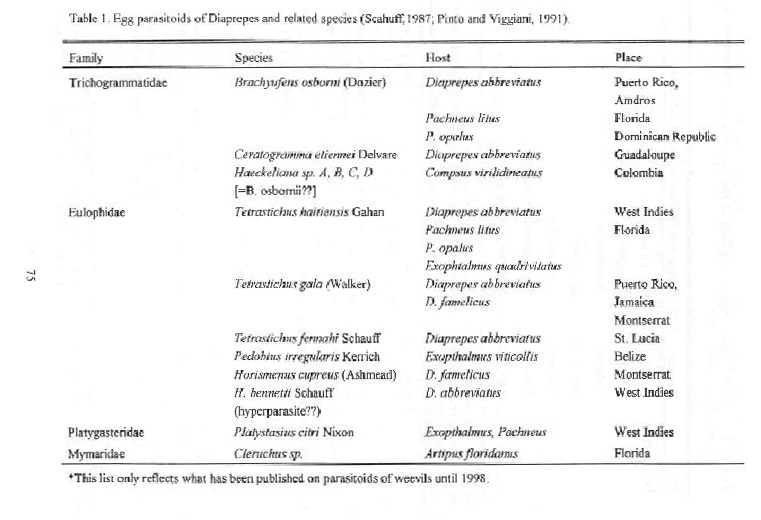
Mated female weevils lay eggs in concealed sites, most often in the space between two adjacent leaves. Egg masses are deposited in a gelatinous cement which seals the leaves together and thus provides protection to the eggs (Browning et al., 1995). Diaprepes eggs are laid on grasses, palm fronds (Peña et al., unpublished data), broad leaf plants (Simpson et al., 1996). No indigenous egg parasitoids of Diaprepes have been observed in Florida (Hall et al., unpublished data). Several egg parasitoids (Brachyufens osborni, Ceratogramma etiennei, Quadrastichus haitiensis, Aprostocetus vaquitarum, Tetrastichus fenahi) are reported from the Caribbean Region, but there are still some doubts of the validity of some reports.
How Does an Egg Parasitoid Locate Diaprepes Eggs?
The adult female parasitoid wasp may use chemical and visual clues to locate eggs. The chemical cues called kairomones, could be on the Diaprepes weevil scales left near the egg by the female weevil during oviposition, on weevil feces or on the sticky substance that the weevil uses to cement eggs between leaves. Some egg parasitoids may also encounter eggs by following sexual pheromones of the weevil. Visual cues (physical configuration of hosts) are probably less used to locate Diaprepes eggs than chemical cues since Diaprepes eggs are laid in concealed sites. Once the parasitoid finds the egg, the parasitoid may drill a hole through the leaf, or it may drill a hole through the egg chorion (egg shell), inserting an egg into the Diaprepes egg (Amalin, unpublished data). Host finding behavior of Ceratogramma etiennei was observed using a time-lapse video camera. C. etiennei walks on the leaves for a few minutes before locating with the antenna the Diaprepes eggs laid between leaves. Once the egg mass is located, C. etiennei drills a hole through the leaf and through the egg chorion and inserts an egg into a Diaprepes egg (Amalin et al., unpublished data).
Diaprepes eggs in the early stages of development are more suitable for parasite development (Amalin et al., unpublished data) than older Diaprepes eggs; especially those in which the head capsule of the larva is visible, are not usually parasitized and if they are, parasitoid survival is much lower (Amalin et al., unpublished data). Since Diaprepes eggs will hatch between 6-10 days after deposition, the parasitoid egg would hatch fast, and the parasitoid larvae would develop quickly. Parasitoid larvae then transform to an inactive pupal stage. After some days, the adult wasp emerges from the pupae and escape the Diaprepes egg by chewing an irregular almost circular hole in the egg shell.
Native Egg Parasitoids of Diaprepes
While several parasitoids, such as Brachyufens osborni are found parasitizing other weevil eggs (blue green weevils, Pachnaeus sp.) in Florida, no native parasitoids of Diaprepes eggs have been detected until now (Hall et al., unpublished data).
Larval and Pupal Parasitoids
After eggs hatch, Diaprepes larvae drops to the soil surface, then proceeds to burrow into the soil to feed on roots. After several months to a year, the larvae will pupate in the soil, close to the soil surface. Even though no Hymenoptera parasitoids of Diaprepes larvae or pupae are identified until now, some insects are parasitoids of larvae or pupae of Coleoptera in the soil. For instance, some Hymenopteran larval and pupal parasitoids of beetles and weevils deposit an egg close to the site where both larvae or pupae are located, or they parasitize the weevil egg and emerge from the weevil pupae. A close example of the behavior of an egg-pupal parasitoid of a weevil can be given with the parasitoid Tetrastichus asparagi Cwfd, a parasitoid of the asparagus beetle. T. asparagi places its eggs in those of the asparagus beetle, and maturity is reached only after the host larva has completed its feeding and formed the pupation cell in the soil. Other parasitoids (i.e., Scollidae wasps) (Table 2) of Coleoptera larvae in the soil, locate host grubs for oviposition, the wasp female must burrow extensively in the soil, and she doubtless often utilizes the burrows that the grubs themselves made. During their feeding period, the Scarabaeid grubs are usually only a few inches beneath the surface and are located without great difficulty; however, females of the wasp Cosila chilensis penetrate to a depth of 40 cm. Females regularly sting the grub, after the grub is paralyzed, the wasp burrows deep into the soil, dragging the grub behind it, and, at a depth of 25 cm to as much as 1 m, a cell is formed within which the grub is placed and the egg then deposited. The depth to which the parasite penetrates with its victim depends largely upon the nature of the soil; in sandy places the parasitized grubs are found at a greater depth than in clay or loam (Clausen, 1940). Other parasitoids that may attack larvae and pupae in the soil or even weevil adults belong in the family Tachinidae. The Tachinidae stand out amongst the dipterous families that are parasitoids of larvae of Lepidoptera, Coleoptera, but some species attack adult bugs, earwigs and grasshoppers (DeBach, 1974). Tachinidae are almost without exception solitary endoparasites and their host does not dies until the larval development of the parasitoid is completed; just occasionally it survives long enough to lay eggs. Adult tachinids are dull-colored flies of moderate size, very bristly but otherwise often resembling houseflies in appearance. They do not have an ovipositor like parasitic Hymenoptera, but they are able to inject their offspring into the body of the host and they have devised other means of gaining entry into the host's body. For example, some species laid their eggs on or in their host, others, laid the eggs on vegetation or on the soil surface, others laid larvae on or in the host, or lay larvae on the soil surface. When a Tachinid larvae is fully grown, it usually emerges from the remains of its host (DeBach, 1974).
Adult Parasitoids
Adult parasitoids have been collected from different beetles and weevils. For instance, the braconid wasps, Microtonus aethiops, Microtonus vittatae oviposit diminutive eggs in the thoracic regions of beetles. These eggs increase in volume by approximately 1200 times during incubation. The parasitoid larvae that develops feeds on the host adipose tissue, sometimes castrating females and male hosts. Full grown parasitoid larvae force their way out of the caudal end of their hosts, invariably killing the beetle. (Smith and Patterson, 1950; Drea, 1968). One adult parasitoid (Microctonus sp.) has been collected by Bullock (1984) from Artipus floridanus weevils in Florida. However, to our knowledge, no adult parasitoids have been collected until today, from Diaprepes in Florida or in the Caribbean Region. One way to determine if adult parasitoids are present would be to the collect weevil adultsfrom infested areas, dissect them and or keep them in confinement for several months until they die. If a parasitoid is the cause for mortality of the weevil, the number of parasitoids emerging per adult weevil should be recorded.
Original Geographical Distribution of Diaprepes Species
Most of the Diaprepesweevil species are found in the Caribbean Region. Diaprepes (Coleoptera: Curculionidae) comprises at least 19 species, of which the better known are D. abbreviatus and D. flamelicus. Therefore, in those areas, where Diaprepes weevils or related species are not considered an economic pest, surveys for natural enemies (i.e., parasitoids) should be initiated. It is expected that in those areas of Diaprepes, low weevil densities have been kept under non-economic levels by effective natural enemies. If the search for parasitoids in the area of origin of the pest fails, it would be important to look for parasitoids of closely related weevil genera that have important biocontrol agents.
Host Plants of Diaprepes abbreviatus
Diaprepes abbreviatus is a highly polyphagous insect that has been found in association with more than 270 plant species (Simpson et al., 1996). Most of these plant species have been introduced into the new world and they have been planted as monocultures (sugarcane, citrus) in different areas of the Caribbean Region. Some parasitoids of Diaprepes may have co-evolved searching for weevils on some native plant species and may stay in their original habitat, while other parasitoids would move from one habitat (wild) to another (citrus, sugarcane) without major problems. Nevertheless, in the case of egg parasitoids, the leaf thickness, leaf pubescence and plant attractants may influence the success of parasitism.
Classical Biological Control of Diaprepes abbreviatus
The introduction and establishment of exotic species for the long term suppression of pests is conveniently referred to as "classical biological control" (Greathead, 1986). Classical biological control is intuitively appealing as a pest management tactic because it involves natural selected components of the ecosystem, is nontoxic and often is self-sustaining. In theory and practice, the goal of classical biological control is to reunite natural enemies with pest species that have invaded new geographical areas with the expectation that effective pest suppression will result (Nechols and Kauffman, 1992). In the case of Diaprepes the objective of this approach is to detect, select and import natural enemies showing promise as biological control agents in the original or adapted ecosystems of Diaprepes abbreviatus or related species. The first question to ask is what have been the previous attempts at biological control of this species. Beavers and Selhime (1975) reported that 6,000 Tetrastichus (= Quadrastichus) haitiensis were released between Apopka and West Palm Beach. These authors reported that two possible reasons for failure to establish this parasitoid were the cold winter temperatures and the lack of correlation between T. haitiensis and D. abbreviatus life cycles in Florida. Also, they observed that competition between B. osborni and T. haitiensis may be detrimental to establishment of T. haitiensis. Earlier Sutton et al. (1972) claimed that one of the problems for establishment of T. haitiensis in Florida was the common application of carbaryl for control of adult weevils. Wolcott (1948) reported that in Puerto Rico during June, T. haitiensis is capable of reducing D. abbreviatus densities. Armstrong (1987) reported that the average parasitism of T. haitiensis in Puerto Rico was 36% between September and December 1980. In Dominica,Whitewell (1991) reported 96% parasitism by an unidentified parasitoid in one egg mass of Diaprepes famelicus. However, one parasitoid species that has been identified from Dominica is Tetrastichus marylandensis Gir. which is present in the USA. Delvare (1988) reported the parasitoid Ceratogramma etiennei (Hymenoptera: Trichogrammatidae) from Guadeloupe. Etienne et al. (1991) developed rearing techniques in the laboratory and determined its life cycle. The life cycle varied from 15 to 30 days with an average fecundity of 19. It can develop in other two Diaprepes species. In absence of chemical treatments the activity of the parasitoid is not negligible (Etienne et al., 1990).
Different steps could be considered when attempting a biological control programme (Zwolfer et al., 1975). They include determining the taxonomic position, life history and geographical distribution of Diaprepes abbreviatus, the current distribution of the pest and close genera or species, assessing host plants in agricultural systems vs. host plants in native systems and defining other climatic regions with the same species or with close related species where exploration for a candidate could occur (Zwolfer et al., 1975):
- Accumulation of Available Information
- Taxonomic position, life history and geographical distribution of Diaprepes abbreviatus.
- Current distribution of the pest and close genera or species.
- Host plants in agricultural systems vs. host plants in native systems.
- Define the center of origin of the species and coexistence of related species.
- Define other climatic regions with the same species or with closely related species where exploration for a candidate could occur.
- Available records of natural enemies.
- Selection of Target Organisms and Exploration Area
- At least two target organisms, i.e., Ceratogramma etiennei and Quadrastichus haitiensis have shown promise in one of the infested areas (Guadeloupe and Puerto Rico).
- Other target organisms, i.e., Quadrastichus haitiensis were released in Florida, but were found to be not efficacious in Central Florida during the 1970s. Again, temperatures and other factors may be affecting the survival of this parasitoid. Currently, after additional releases in 1999, these parasitoids have been recovered from different areas of Florida (Hall, unpublished data).
- Select areas where closely related species have been observed and where the pest species are kept under non-economic levels.
- Exploration Areas
- Search should be initially undertaken in areas where the pest is native and where a rich diversified complex of natural enemies exists, as some could be host-specific. However, this aspect should not blind the researcher for the possibility that suitable control agents exist outside the native areas of the host.
- Climate and environmental conditions similar to those of the country where introductions are intended.
- Faunal and faunistic history and habitat structures similar or diverse in the area of exploration?
- Inventory Research and Investigations on Selected Species
- Criteria for selection of suitable control agent.
- Localities with low insect density; concentrate or present different host stages in the field; then collect host material and parasitoids of host material.
- Determine presence or absence of hyperparasitoids.
- Are these natural enemies found in other climates? areas? Are these natural enemies found parasitizing the host pest on different plant species?
- Importation
- The selection for parasitoids must be assisted by a systematist. Systematic research has long been recognized as vital ingredient of the formula for success in biological control programs. If a project is not supported by a systematist, lack of adequate, timely and accurate information has caused the delay and failure of many biological control programs (Schauff, 1992).
- Host specificity is a highly desirable attribute of arthropod natural enemies imported for biological control of exotic pests. Therefore, whenever possible, candidates for biological control should be collected or reared from the target pest species or a closely related species. Natural enemies should also be collected from the native home or source of the pest invader, from an ecologically homologous area (Nechols et al., 1992). Thus, when evaluating new natural enemies for importation or release, it is important to determine whether they are likely, or able, to feed on valued non target species. For instance Ceratogramma etiennei was tested for specificity, against lepidopteran eggs of native butterfly species, weevils released for biological control of weeds and closely related weevils (i.e., Pachnaeus sp.). C. etiennei only accepted Pachnaeus sp., as an additional host.
- Establish subcontracts for entomologists in the area to send material to researcher.
- Rearing of pest in area of introduction.
- Type of Releases
- Augmentation is the periodic release of a natural enemy that does not occur naturally in sufficient numbers to keep a pest below damaging levels. Augmentation can be carried out by inundative releases or inoculative releases. The inundative approach is achieved by flooding the crop with multiple releases. In this case, the released insects control pests at the present time, but there is little expectation that later generations will persist at sufficient levels to provide control. This approach requires a large number of natural enemies at a precise time when egg pests are present. Correct timing requires coordination between the rearing facility and field staff.
- Inoculative releases involve one or several releases to establish populations of the natural enemy before pest densities have begun to increase. The natural enemy reproduces on the target pest or an alternate host and its population increases to levels sufficient to control the target pest later in the season.
- Conservation
- Conservation as a biological control method includes crop management practices that protect and encourage natural enemies and increase their impact on pests. Examples include using only selective pesticides and planting strip crops around fields to provide food and habitat for natural enemies. Ceratogramma etiennei has been commonly found parasitizing Diaprepes eggs on palm leaf fronds (Amalin et al., unpublished data). During the last 2 years, we have introduced from Guadeloupe, cultured and released, a parasitic wasp, Ceratogramma etiennei (Hymenoptera: Trichogrammatidae) which is highly specific to eggs of the genus Diaprepes (Etienne et al., 1990; Peña et al., 1998). During the last year, the parasitoid is considered established, at least in southern Florida. Despite of the promise of this biological control agent, mechanisms for release methodologies, release rates and evaluation strategies need to be investigated and improved in order to obtain a successful establishment of the parasite in the field.
References
1. Adair, R. C., H. Nigg, S. Simpson, and L. Lefebre. 1998. Ovipositional preferences of Diaprepes abbreviatus (Coleoptera: Curculionidae). Florida Entomol. 81:225-234.
2. Adair, R. C. 1994. A four-year field trial of entomopathogenic nematodes for control of Diaprepes abbreviatus in a flatwoods citrus grove. Proc. Fla. State Hort. Soc. 107:63-68.
3. Anonymous. 1995. Nematodes play increased role in controlling weevils. Florida Grower and Rancher, September, 88:6-7.
4. Anonymous. 1996. Detections of Diaprepes abbreviatus in Florida. Fla. Dep. Agric. Cons. Serv. Div. Plant Ind. Bureau of Eradication and Control. 2 p.
5. Armstrong, A. 1987. Parasitism of Tetrastichus haitiensis Gahan on egg masses of Diaprepes abbreviatus in Puerto Rico. J. Agric. Univ. P. R. 71:407-409.
6. Beavers, J. B. and A. G. Selhime. 1975. Further attempts to establish the weevil egg parasite, Tetrastichus haitiensis in Florida. Fla. Entomol. 58:29-31.
7. Beavers, J. B., S. A. Lovestrand, and A.G. Selhime. 1980. Establishment of the exotic parasite Tetrastichus haitiensis (Hym: Eulophidae) and recovery of a new Trichogramma (Hym: Trichogrammatidae) from root weevil egg masses in Florida. Entomophaga 25:91-94.
8. Beavers, J. B. and W. Schroeder. 1980. Sugarcane rootstalk borer weevil, Diaprepes abbreviatus; insect growth regulators. Insecticide and Acaricide Tests. 5:234.
9. Browning, H. W., R. McGovern, L. Jackson, D. Calvert and W. Wardowski. 1995. Florida Citrus Diagnostic Guide. Florida Science Source, Lake Alfred, FL. 244 p.
10. Bullock, R.C. 1984. Endoparasitic braconid (Hymenoptera: Braconidae) attacks little leaf notcher Artipus floridanus Horn (Coleoptera: Curculionidae) in Florida. Florida Entomol. 67:571-572.
11. Bullock, R. C., C. McCoy and J. Fojtik. 1988. Foliar sprays to control adults of the citrus root weevil complex in Florida. Proc. Fla. Hort. Soc. 101:1-5.
12. Clausen, 1940. Entomophagous Insects. McGraw Hill, New York. 688 p.
13. DeBach, P. 1974. Biological control by natural enemies. Cambridge University Press, New York., 323 p.
14. Delvare, G. 1988. Ceratogramma etiennei n. sp. , Parasite a la Guadeloupe de Diaprepes abbreviatus L. (Hymneoptera: Trichogrammatidae; Coleoptera: Curculionidae). Revue fr. Ent. 10:1-4.
15. Drea, J. 1968. Castration of male alfalfa weevils by Microtonus spp. J. Econ. Entomol. 61:1291-1295.
16. Etienne, J., H. Mauleon and B. Pinturau. 1990. Biologie et dynamiquex de Ceratogramma etiennei (Hymneoptera: Trichogrammatidae) parasite de Diaprepes abbreviatus (L.) (Coleoptera: Curculionidae) en Guadeloupe. Les Colloques de L INRA 58: 459-68.
17. Fennah, R. G. 1942. The citrus pests investigation in the Windward and Leeward Islands British West Indies 1937-1942. Agric. Advisory Dept. Imperial College of Trop. Agric. Trinidad, B. W. I., 66 pp.
18. Hall, D. G. 1995. A revision to the bibliography of the sugarcane rootstalk borer weevil, Diaprepes abbreviatus (Coleoptera: Curculionidae) Florida Entomol. 78:364-377.
19. Jaffe, K., H. Mauleon, and A. Kermarrec. 1991. Qualitative evaluation of ants as biological control agents with special reference to predators on Diaprepes sp. (Coleoptera: Curculionidae) on citrus groves in Martinique and Guadeloupe. Recontres Caraibes en Lutte Biologique, Guadeloupe. INRA, pp. 405-416.
20. McCoy, C., and S. E. Simpson. 1994. Past and current IPM strategies to combat the spread of Diaprepes abbreviatus in Florida citrus. Proc. Caribbean Food Crops Soc., St. Thomas 30:247-255.
21. McCoy, C. W., E. Quintela, S. Simpson and J. Fojtik. 1995. Effect of surface-applied and soil-incorporated insecticides for the control of neonate larvae of Diaprepes abbreviatus in container-grown citrus. Proc. Fla. Hort. Soc. 108:130-136.
22. Nechols, J. R., and W. C. Kauffman. 1992. Overview. In: W. Kauffman and J. Nechols, eds., Selection criteria and ecological consequences of importing natural enemies. Thomas Say Publications in Entomology: Proceedings. pp. 1-5.
23. Nechols, J. R., W. C. Kauffman and P. W. Schaefer. 1992. Significance of host specificity in classical biological control. pp. 41-52. In: W. Kauffman and J. Nechols, eds., Selection criteria and ecological consequences of importing natural enemies. Thomas Say Publications in Entomology: Proceedings.
24. Pavis, C., C. Malosse, P. Ducrot, F. Howse, K. Jaffe and C. Descoins. 1992. Defensive secretion of first instar larvae of rootstalk borer weevil, Diaprepes abbreviatus (Coleoptera: Curculionidae), to the fire-ant Solenopsis geminata (Hymenoptera: Formicidae). J. Chemical Ecol. 18:2055-2068.
25. Peña, J. E., Etienne, J., Duncan, R., and Pinto, J. 1998. Introduction of Ceratogramma etiennei (Hymenoptera: Trichogrammatidae) for biological control of Diaprepes abbreviatus in Florida, USA. In Egg Parasitoids, 5th International Symposium, IIOBC, Cali, Colombia, March, 1998, S. A. Hassan, ed. pp.145-148.
26. Pinto, D. J. and G. Viggiani. 1991. A taxonomic study of the genus Ceratogramma (Hymenoptera: Trichogrammatidae). Proc. Entomol. Soc. Washington 93:719-732.
27. Quintela, E., and C. W. McCoy. 1997. Effects of inidachloprid on development, mobility, and survival of first instars of Diaprepes abbreviatus (Coleoptera: Curculionidae). J. Econ. Entomol. 90:988-995.
28. Schauff, M. E. 1987. Taxonomy and identification of the egg parasites (Hymenoptera: Platygasteridae, Trichogrammatidae, Mymaridae and Eulophidae) of Citrus weevils (Coleoptera: Curculionidae) Proc. Entomol. Soc. Washington 89: 31-42.
29. Schauff, M. E. 1992. Systematics in biological control. pp. 15-25. In: W. Kauffman and J. Nechols, eds., Selection criteria and ecological consequences of importing natural enemies. Thomas Say Publications in Entomology: Proceedings.
30. Schroeder, W. J. and J. B. Beavers. 1977. Citrus root weevils in Florida: Identification, biology and control. Proc. Intern. Soc. Citriculture 2:498-500.
31. Schroeder, W. J. and D. Green. 1983. Diaprepes abbreviatus: oil sprays as a regulatory treatment, effect on egg attachment on Dracaena marginata. J. Econ. Entomol. 76:1395-1396.
32. Schroeder, W.1990. Suppression of Diaprepes abbreviatus (Coleoptera: Curculionidae) adult emergence with soil application of entomopathogenic nematodes (Nematoda: Rhabditida). Fla. Entomol. 73:680-683.
33. Schroeder, W. 1996. Diflubenzuron residue: reduction of Diaprepes abbreviatus (Coleoptera: Curculionidae) neonates. Florida Entomol. 79:462-463
34. Schroeder, W. J., and P J Sieburth. 1997. Impact of surfactants on control of the root weevil Diaprepes abbreviatus larvae with Steinernema riobravis. J. Nematology. 29:216-219.
35. Simpson S.E., H. Nigg, N. Coile and R. Adair. 1996. Diaprepes abbreviatus (Coleoptera: Curculionidae): Host plant associations. Environ. Entomol. 25:333-349.
36. Smith, O., and A. Patterson. 1950. Microtonus vittatae, a parasite of adult flea beetles, and observations on hosts. J. Econ. Entomol. 43:581- 585.
37. Sutton, R.A., G. Selhime and W. McCloud. 1972. Colonization and release of Tetrastichus haitienisis as a biological control agent for citrus root weevils. J. Econ. Entomol. 65:184-185.
38. Thompson, W. 1943. A catalogue of the parasites and predators of insect pests. Section 1. Parasite Host Catalogue, Belleville, Ontario. p. 41.
39. Sutton, R. A., G. Selhime and W. McCloud. 1972. Colonization and release of Tetrastichus haitienisis as a biological control agent for citrus root weevils. J. Econ. Entomol. 65:184-185.
40. Thompson, W. 1943. A catalogue of the parasites and predators of insect pests. Section 1. Parasite Host Catalogue, Belleville, Ontario. p. 41.
41. Tryon, H. 1986. The stripped earwing and ant predators of sugarcane rootstock borer in Florida Citrus Fla. Entomol. 69:336-343.
42. Whitcomb, W. H., T. Gowan, and W. Buren. 1982. Predators of Diaprepes abbreviatus larvae Sugarcane rootstalk borer weevil in Florida. Florida Entomol. 65:150-158.
43. Whitwell, A. C. 1990. Diaprepes spp. problems in Dominica and some possible solutions. pp. 529-541, In, recontres Caraibbes en lutte biologique, C. Pavis and A. Kermarrec, eds. INRA, Paris. Les colloques no. 58.
44. Wolcott, G. N. 1936. Control of soil inhabiting grubs of Puerto Rico. J. Agric. Univ. P.R. 34:333-337.
45. Wolcott, G.N. 1948. The insects of Puerto Rico: Coleoptera. J. Agric. Univ. Puerto Rico 32:225-416.
46. Zwolfer, H., M. A. Ghani and V. P. Rao. 1975. Foreign exploration and importation of natural enemies, pp. 89-207, In Huffaker, C.B. and P.S. Messenger, eds. Theory and practice of biological control, Academic Press, New York.