Biology of Diaprepes abbreviatus in the Laboratory and Field
Philip A. Stansly
Southwest Florida REC
University of Florida/IFAS
Immokalee, FL
Diaprepes is thought to have originated in the Caribbean where it was known principally as a pest of sugarcane, but also of citrus and other crops. Not surprisingly, the earliest biological studies of D. abbreviatus were conducted in the Caribbean, most notably by G. N. Wolcott (1933, 1936) in Puerto Rico. Focus shifted to Florida after the weevil was detected in 1964 (Woodruff, 1964), with studies of, Beavers and Selhime (1975a), Beavers (1982), and later Quintela et al., 1998, Rogers et al., 2000. Results from all these authors were consistent in reporting long and variable development time and low survivorship in the laboratory. Beavers (1982) even thought that prolonged larval development would "preclude mass rearing". However, more recent studies (Lapointe and Shapiro, 1999; Lapointe, 2000) have shown, that time and variation can be reduced by holding constant temperature and moisture content of the artificial diet substrate. This result will markedly increase the efficiency of rearing facilities. Still to be clarified is the way in which temperature, humidity, host plant quality and possibly other factors interact in the field to determine the observed annual patterns of abundance.
Adult Longevity
Wolcott (1936) gave 160 and 123 days as mean longevity of 13 and 16 females and males respectively, with at least one living up to 336 days. This interval included time in a quiescent period between eclosion from the pupa and emergence from the soil. The actual time above ground was quite a bit shorter especially for the male (less than 2 months) compared to the female (over 4 months). He reported 3 to 7-day preoviposition period between female emergence from the soil and initiation of egg laying. Beavers (1982) reported female longevity on artificial diet as 147 ± 17 days compared to 135 ± 22 for males. He mentioned no quiescent period but observed a 22 ± 3 day preoviposition period following eclosion from the pupa. The discrepancy in preoviposition periods is evidently due to the greater maturity of Wolcott's weevils as they emerged from the ground compared to Beavers (1982) as they eclosed from the pupae.
Oviposition
Eggs are normally deposited in the field as masses or clusters sandwiched between two leaves. Eggs are laid on sugarcane and other grasses between the split tips of an individual leaf. The sandwich is cemented together by a gelatinous adhesive secreted by the female at the time of oviposition. Most (80%) eggs were deposited between the hours of 6 PM and 6 AM during August in Florida (Schroeder, 1981). Paper strips can also be used as an oviposition substrate and are preferred to leaves (Wolcott, 1933), especially waxed or freezer paper. The number of eggs per cluster laid on oiled paper in the laboratory was reported by Wolcott (1936) from 9 individuals as ranging from 30 to 264 with an average of 52 eggs per day over 97 days or 5070 eggs per female. Beavers (1982) reported (mean ± standard error) 6517 ± 931 over 125 days (longevity minus preoviposition period) or 52 eggs/day. He observed an average 69 ± 8 eggs per egg mass and a hatch rate of 89% at 25°C and 80% RH. Most (90%) eggs hatched between 7 and 8 days under these conditions.
Larval Development
Neonate larvae escape the leaf sandwich by wiggling around or through the adhesive secretion and falling to the ground. They penetrate the soil surface through a tiny crack or crevice and seek a rootlet to feed upon (Wolcott, 1936). Beavers (1982) observed that of over 1000 neonates maintained without food but with adequate moisture, all were alive after 25 days and all did not die until 60 days. Wolcott (1933, 1936), used a corn or lima beans to maintain at least 240 larvae which he followed individually. Only 3 of these survived from egg hatch to adult emergence which took an average of 236 days. He reported an average 57 days necessary for the first 6 instars, but with considerable seasonal variation and more individual variation. He believed there were usually 8 instars but observed as few as 6 or as many as 16, although most growth occurred during the first 6 instars. At the end of this growth period, he observed the larva to persist but not grow for a period of from 55 to 338 days. This so called "obligitory diapause" lasted from 2 to 13 months and was independent of season, soil humidity or temperature (Wolcott, 1933).
Beavers (1982) reported most growth on artificial media during the first 3 to 4 months when larvae reached a maximum weight of about 550 mg. Larval development time for diet-reared females (N = 344) and males (N = 156) was 377 ± 4.8 and 378 ± 6.4 days respectively. These results indicated a long but relatively constant larval period as indicated by coefficients of variation (standard deviation/mean) of 23.6 and 21.1 respectively. Possibly, their diet was drying out at a constant rate. According to Quintela et al. (1998), there are 10 or 11 larval instars although most weight gain on diet occurred in their studies through the 8th instar. They reported an average 153 ± 28 days to pupation (CE = 108%) of which all but 58 ± 16 were spent in the last 3 instars with little feeding or growth. Poor diet quality was blamed for few larvae reaching pupation and could have been the cause of the prolonged and variable late larval stage. However, mean larval weight on diet (555 mg) was 4 times greater than from potted citrus in the greenhouse (139 mg) with larvae from citrus roots in the field intermediate (297 mg). Rogers et al. (2000) reported similar results from potted citrus, with most weight gain occurring in the first 70 days. Apparently, potted citrus provides inadequate nutrition and even citrus in the field is deficient over time compared to artificial diet.
With the exception of Wolcott's experiments, all the above trials were conducted at constant temperatures ranging from 25 to 27 ± 2° C. Lapointe and Shapiro (1999) ran their experiments in controlled temperature cabinets at 25 ± 1° C but focused on substrate moisture as a variable. Larvae were reared on diet through 68 days, then transferred to soil with moisture content between 20 and 80%. Highest mortality was observed at the two moisture extremes, although there was no difference in days to pupation among moisture treatments. During another series of experiments to test temperature effects (Lapointe, 2000), care was taken to maintain constant moisture in the diet by sealing cups in plastic bags and replacing diet every 28 days. Growth rate was found greatest at 30°C during the first 14 days after hatching. However, weight and survivorship were lower and development was prolonged for later instars at 30°C compared to 26°C, the optimal temperature tested. Under these conditions, larval development was only 106 and 125 days (CE = 13% and 15%) in the two experiments reported. Thus, if a diapause period does occur at the end of larval growth, it is not obligatory.
Pupation and Adult Emergence
When ready to pupate the larva forms a vertical chamber in the soil and rests in it with its head up (Wolcott, 1936). Soil forming the wall of the chamber is compacted by active movements of the larva. Pupation occurs within 2 or 3 week after the chamber is formed. This author observed the pupal period to vary between 17 and 26 days but thought that "slightly more than 2 weeks" was normal. Beavers (1982) reported a pupal period of 15.2 ± 0.43 and 15.4±0.77 days from females and males respectively. Quintela et al. reported 17±3.8 days at 27°C. Lapointe (2000) reported the pupal stage ranging from 32 or 35 days at 22°C to 16 or 15 days at 30°C. Thus, there seems to be little variability in pupal development time under constant conditions in the laboratory.
When is the larva ready to pupate? Schroeder (1987) noted that 86% of larvae removed from diet at 120 days and placed in soil at 27°C pupated. Average time to pupation was 20 ± 4 days and average time to adult emergence was 49 ± 3 days after placement in the soil. In contrast, only larvae placed in diet under the same conditions pupated. These results would indicate that pupation can be induced in fully grown larvae by removing the food source.
Wolcott (1936) observed that the adult remained "quietly" in its pupal chamber long after appearing physically able to emerge. He reported a minimum of 11 days and a maximum of 126 days for this quiescent period, with over half the observed adults emerging between 33 and 48 days. However, if the larva were disturbed it would no longer remain in the cell of its own accord. Inability to induce emergence of adults from pupal chambers in soil tube with water or temperature changes led him to believe that external factors of temperature or moisture had no effect on Puerto Rican Diaprepes. Apparently, the quiescent period for diet-reared weevils in the laboratory was incorporated into the prolonged pre-oviposition period observed by Beavers (1982).
Life Cycle Summary
In attempting to summarize data from these different sources we find considerable differences albeit with a trend toward shorter life cycles in more recent laboratory studies (Table 1). These later studies are characterized by more constant rearing conditions, particularly Lapointe and Shapiro (1999) and Lapointe (2000). Thus, it appears that under optimal conditions of temperature, humidity and nutrition, the life cycle can be completed in little more than 5 months.
Phenology
Wolcott (1936) believed the "normal, original lifecycle" of D. abbreviatus to be one year, with adults appearing above ground in late spring and early summer. The presence of adults year-round was attributed to variation caused by the "obligatory" diapause period of the late instar larva and the quiescent period of the adults prior to emergence. He thought this variability to be an adaptation for avoiding attack by the egg parasitoid Quadrasticus haitiensis that was most abundant in early summer. In support of this notion, he cited reports from Barbados where the parasitoid was not present and (consequently) adults appeared suddenly in response to "spring rains".
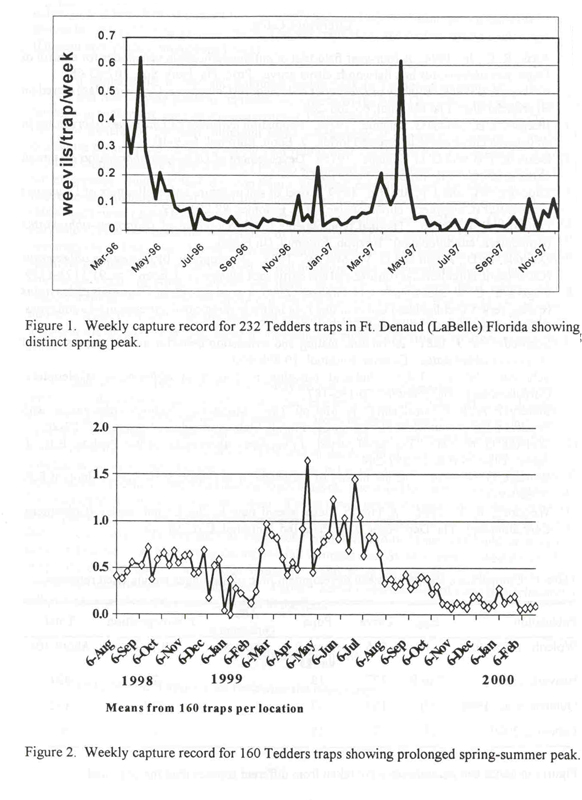
Both similar and very different patterns have been observed in Florida. Beavers and Selhime (1975b) monitoring trees visually, observed highest populations in late summer or early fall and speculated that emergence was driven by summer rains. However, the observed trend may have reflected an earlier emergence peak that was not detected due to the method of sampling adults in trees. Stansly et al. (1997) monitoring with pyramidal Tedders traps near LaBelle, Florida, reported a sharp and distinct emergence peak at the end of April, followed by declining capture during summer and a weak, secondary peak in late fall (Fig. 1). The same pattern existed in 2 other nearby groves (unpublished data). However, in a 4th grove, still in the vicinity, about 4 times more weevils are captured routinely and the peak has been prolonged well into summer (Fig. 2). Weevils were detected most recently in this grove (1996) possibly indicating a recent infestation. Adair (1994) using cone emergence traps near Vero Beach, also observed peaks in late spring over 4 years, although in a less defined pattern than generally seen in LaBelle. Emergence patterns at several groves in central Florida as indicated by Tedders traps show the typical strong peak in late spring (Duncan and McCoy, unpublished data).
It appears that a clear and dominant spring peak in Tedders trap captures is the norm in Florida and that varying patterns represent exceptions to the rule. Given that this salient feature of the capture cycle represents emergence patterns, Diaprepes must be largely univoltine in Florida, i.e. have one generation per year, although some overlapping of generations occurs. Clearly then, conditions are not ideal for development year round since the weevil has the potential to reproduce in 5 to 6 months. Beavers and Selhime (1975b) suggested soil moisture as an environmental trigger for emergence, implying that dry conditions inhibited completion of development. However, Lapointe and Shapiro (1999) found that older larvae were adversely affected only by the highest (80%) and lowest (20%) soil moisture regimes tested and pupae were unaffected. On the other hand, Lapointe (2000) mentions the importance of maintaining proper moisture levels in diet for larval development and survival. Therefore, a distinction should be drawn between moisture in the larval substrate and moisture in the larval food. If soil moisture were a controlling factor, why would peak emergence occur in April, normally the driest month in Florida with least rainfall and high evapotranspiration? Apparently, larvae can tolerate low soil moisture if they can obtain adequate moisture from their diet. In any case, the irrigation systems typically utilized in Florida citrus, including the monitored groves, serve to maintain soil moisture relatively constant within the root zone.
Soil temperatures in the shade probably do not become unfavorably hot for long periods of time in Florida, but may become unfavorably cool in winter (Lapointe, 2000), slowing development until spring. Food quality and quantity would also likely be poor during the winter months as citrus trees tend to become dormant, and improve after the spring flush when nutrients again flow from foliage to roots. These factors would slow weevil development during winter and speed it to a conclusion in the spring. On the other hand, a second generation of weevils emerging in the fall would face unfavorable conditions leading to poor development and low survivorship. Adults would find a paucity of young flush to feed upon. Young larvae could starve for lack of active roots or development slowly in response to cool temperatures near the soil surface. Finally, there is evidence that natural mortality factors such as indigenous entomophagous nematodes build up during the summer and occur at high levels during the fall (Duncan and Stansly, unpublished data). Thus, temperature food quality and biotic mortality could work in concert to synchronize weevil populations into the yearly cycle observed. Perhaps better understanding of these interrelationships will provide useful management strategies in the future.
Acknowledgement
Thanks to S. L. Lapointe for reviewing the manuscript.
Literature Cited
1. Adair, R. C., Jr. 1994. A four-year field trial of entomopathogenic nematodes for control of Diaprepes abbreviatus in a flatwoods citrus grove. Proc. Fla. Hort. Soc. 107:63-68.
2. Beavers, J. B. 1982. Biology of Diaprepes abbreviatus (Coleoptera: Curculionidae) reared on an artificial diet. Fla. Entomol. 65:263-269
3. Beavers, J. B. and G. G. Selhime. 1975a. Population dynamics of Diaprepes abbreviatus in an isolated citrus grove in central Florida. J. Econ. Entomol. 69:9-10.
4. Beavers, J. B. and G. G. Selhime. 1975b. Development of Diaprepes abbreviatus on potted citrus seedlings. Fla. Entomol. 58:271-273.
5. Lapointe, S. L. and J. P. Shapiro. 1999. Effect of soil moisture on development of Diaprepes abbreviatus (Coleoptera: curculionidae). Fla. Entomol. 82:291-299.
6. Lapointe, S. L. 2000. Thermal requirements for development of Diaprepes abbreviatus (Coleoptera: curculionidae). Environ. Entomol. (In Press).
7. Quintela, E. D., J. Fan and C. W. McCoy. 1998. Development of Diaprepes abbreviatus (Coleoptera: curculionidae) on artificial and citrus root substrates. J. Econ. Ent. 91:1173-1179.
8. Rogers, S., C. W. McCoy and J. H. Graham. 2000. Larval growth of Diaprepes abbreviatus (Coleoptera: Curculionidae) and resulting root injury to three citrus varieties in two soil types. J. Econ. Entomol. 93: (in press).
9. Schroeder, W. J. 1981. Attraction, mating and ovipostion behavior in field populations of Diaprepes abbreviatus. Environ. Entomol. 10:898-900.
10. Schroeder, W. J. 1987. Induced pupation in Diaprepes abbreviatus (Coleoptera: Curculionidae). Fla. Entomol. 70:186-187.
11. Stansly, P. A., R. F. Mizell and C. W. McCoy. 1997. Monitoring Diaprepes abbreviatus with Tedders traps in southwest Florida citrus. Florida State Horticulture Society 110: 22-26.
12. Wolcott, G. N. 1933. The larval period of Diaprepes abbreviatus at Rio Piedras, P.R. J. Agric., Univ. of P.R. 17:257-264.
13. Wolcott, G. N. 1936. The life history of Diaprepes abbreviatus L. J. Agric., Univ. of P.R. 20:883-914.
14. Woodruff, R. E. 1964. A Puerto Rican weevil new to the United States (Coleoptera: Curculionidae). Fla. Dep. Agric. Div. Plant Ind. Entomol. Circ. 30:1-2.
Table 1. Comparative results for mean development time of life stages among cited references.
| Publication | Egg | Larva | Pupa | Adult Quiescence |
Pre-oviposition | Total |
|---|---|---|---|---|---|---|
| Wolcott, 1933/1936 |
7 | 236 | About 2 weeks | About 40 | 3 to 7 | About 303 |
| Beavers, 1982 |
7 to 8 | 377 | 15 | 22 | 421 | |
| Quintela et al., 1998 |
(7) | 153 | 17 | (14) | 192 | |
| Lapointe, 2000 |
(7) | 126 | 15 | 14 | 162 | |
| Figures in italics and parentheses were taken from different sources than the one cited. | ||||||