Entomopathogenic Nematodes as a Component of Citrus Root Weevil IPM
L. W. Duncan, C. W. McCoy, and J. H. Graham
University of Florida, IFAS
Citrus Research and Education Center
Lake Alfred, FL 33850, USA
D. I. Shapiro
USDA, ARS
SE Fruit and Tree Nut Lab
Byron, GA 31008
Abstract
The entomopathogenic nematodes, Steinernema riobrave, Heterorhaditis bacteriophora and H. indicia are routinely used by many citrus growers in Florida as a component of IPM programs to control a root weevil, Diaprepes abbreviatus, because: 1) D. abbreviatus is currently a major biological threat to citriculture in the state; 2) until 1998, no effective chemical pesticides were registered for control of weevil larvae in soil; 3) the short-term efficacy of S. riobrave has been consistently documented; and 4) use of nematodes is relatively inexpensive. Although unexploited, natural control of root weevils by undescribed species of indigenous nematodes in Florida appears to be high. However, major questions remain regarding optimum use of S. riobrave and other entomopathogenic nematodes. Constraints on sampling methodologies have impeded the derivation of economic thresholds and of models of weevil population dynamics that could be used to estimate optimum timing and frequency of nematode treatments. The recommended nematode application rate appears to be adequate for treatment of young trees, but may be too low to provide consistent results in larger mature trees. Research is also needed to estimate: 1) the long-term efficacy of nematodes against weevils; 2) the relative efficacy of nematodes compared to insecticides in an IPM program; and 3) the profitability of D. abbreviatus IPM.
Introduction
Several insect species in the family Curculionidae are commonly referred to as citrus root weevils. In Florida, and throughout the Caribbean region, the West Indian sugarcane rootstalk borer weevil, Diaprepes abbreviatus L., is the root weevil of greatest economic significance to citrus. The insect was first detected in Florida in 1964, and currently infests an estimated 30,000 of the 845,000 acres of commercial citrus orchards. During the past decade, D. abbreviatus has become the most serious biological threat to the well-being of citriculture in Florida because of its high incidence, its devastating effect on trees, and because cost-effective IPM strategies have been elusive. Prior to 1998, attempts to intervene in the soil-borne phase of the weevil life cycle were hampered by the absence of registered, effective soil-applied pesticides, due to environmental concerns. For these reasons, the use of entomopathogenic nematodes to manage citrus root weevils has had a high priority for more than a decade among both researchers and citrus growers in Florida.
Adult D. abbreviatus feed and oviposit on the leaves of citrus and alternate host plants in orchards (Fig. 1). Newly-hatched (neonate) larvae drop to the soil where they develop for 4-9 months while feeding on the root systems of trees. Pupation occurs in the soil. Young larvae feed initially on the small fibrous roots (Fig. 2), but as they increase in size they feed on the cortex of increasingly larger roots. The insects create long lesions or channels in the bark of large roots, which are then infected by the root-rotting fungi Phytophthora nicotianae Dastur, and P. palmivora (Butler) Butler (McCoy, 1999; Graham and Menge, 1999). The interaction between root weevils and plant pathogenic fungi results in one of the most severe decline syndromes affecting citrus. Trees are sometimes killed by a resulting crown rot, but more typically trees decline severely and irreversibly due to cambium girdling and death of large structural roots.
It is often necessary to remove and replant the majority of trees within root weevil-infested orchards. Large scale replanting has serious economic consequences because maximum fruit yield is not attained for 10-15 years. Moreover, weevil-resistant rootstocks are unavailable and the costs of managing weevils in replanted orchards may exceed $250/acre/year. Recommended IPM of D. abbreviatus currently consists of the use of insecticides (e.g. carbaryl) and ovicides (e.g. spray oil or diflubenzuron + spray oil to open adhering leaves that protect egg masses) for above-ground control of eggs and adult insects, fungicides (e.g. metalaxyl) for control of Phytophthora spp., and soil-applied insecticides (imidacloprid or bifenthrin) or entomopathogenic nematodes for control of soil-borne stages of the insect (Knapp, 1998). Increased application frequency of water and fertilizer is also recommended to improve the tolerance of trees to the loss of roots caused by root weevils.
When Steinernema carpocapsae Weiser was reported to have efficacy against D. abbreviatus (Schroeder, 1990), the nematode quickly became commercially available and widely used in Florida orchards. However, perceived failure of the commercial product in the field and subsequent research (Schroeder, 1994; Duncan et al., 1996) resulted in changing the commercially available species from S. carpocapsae to S. riobrave Cabanillas, Poinar & Raulson and H. bacteriophora Poinar, Kanunakar & David. Research during the past 5 years (Table 1) has confirmed the potential effectiveness of S. riobrave as a component in IPM of citrus root weevils (Bullock et al., 1999; Duncan and McCoy, 1996; Downing et al., 1991). Moreover, treatment costs $25 per acre are relatively modest compared to those for most soil-applied chemical pesticides.
Despite a widespread acceptance of entomopathogenic nematodes for use in weevil IPM, a number of concerns exist. Neither researchers nor growers consider current IPM programs to be adequate for economic management of citrus root weevils. There are no published studies of the profitability of these programs and anecdotal evidence suggests that long-term weevil control is variable and in some cases marginal. The research constraints to answering questions of efficacy and profitability are enormous. There are no methods to directly assess population densities of weevils in soil, chemical attractants for adults are unknown, and adult monitoring methods are inefficient. Root loss and root damage cannot be assessed non-destructively. The effect on yield of mitigating root damage is complex, requiring long-term study of several crop cycles. Methodology problems such as these have constrained research to determine the insect life cycle, the incidence and causes of natural control, optimum application timing and rate of entomopathogenic nematodes, root loss-yield relationships, and insect economic thresholds.
However, ongoing research is attempting to address some of these problems in order to provide information needed to optimize the use of tactics for IPM of citrus root weevils. The objectives of this paper are to use data from published and ongoing studies to describe how entomopathogenic nematodes are currently used by citrus growers, and to identify some of the questions that are being studied to improve the utilization of these organisms.
Choice of Nematode Species and Formulation
Several indigenous entomopathogenic nematode species (some undescribed) that parasitize citrus root weevils occur in Florida citrus orchards (Authors and K. Nguyen, unpublished). For example, Heterorhabditis indica was recently described from South Florida and is now available commercially. Other nematodes available commercially for use in Florida citrus are S. riobrave and H. bacteriophora. Each nematode species has different search strategies that affect the horizontal and vertical distance it migrates (Kaya et al., 1993), and each persists differently under different conditions (see below). Similarly, recent laboratory data indicate that H. indica may have higher virulence than other species against younger (ca. 4th instar) D. abbreviatus larvae (Shapiro et al., 1999), whereas other experiments indicate S. riobrave is more virulent against older (7-11th instar) larvae (Shapiro and McCoy, unpublished). Published research suggests that S. riobrave performs as well as or better than other available species under conditions tested to date (Table 1). However, it should be noted that most studies have reported results of short-term evaluations, and did not measure long-term efficacy that could result from superior persistence in soil of a particular species. Similarly, further evaluation of indigenous nematode species may reveal characteristics that are advantageous compared to commercially available species.
The quality of formulated nematodes is also important when choosing a commercial product. Entomopathogenic nematodes differ in their ability to remain viable when commercially formulated and quality control can vary among products and production batches. Nematodes in liquid formulation cannot be stored by the grower for more than 2-3 days and their viability is generally evaluated just prior to shipment. Alternatively, when using a granular formulation, it is advisable for the user to have a means to evaluate the motility of nematodes just prior to use. The proportion of motile nematodes is useful to estimate viability which sometimes deteriorates markedly during permitted storage times as long as 1 month.
Application Timing And Frequency
Recommendations about when and how often to apply entomopathogenic nematodes have been inferred from seasonality of emergence of adult insects from the soil, from estimates of nematode persistence following application, from research on physical causes of nematode mortality, and by considering the cost of applying nematodes. However, significant gaps exist in our understanding of the insect population dynamics and the spatial/temporal relationships between nematode density and efficacy.
Nematode Persistence and Natural Control by Nematodes
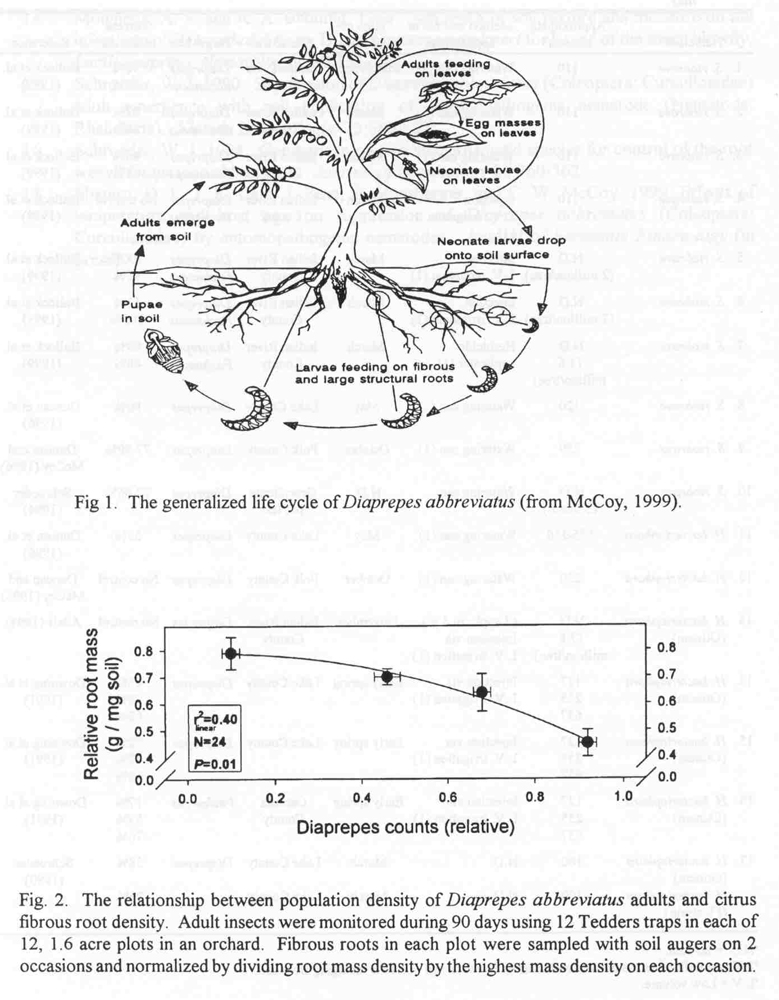
When entomopathogenic nematodes are applied to soil in Florida, their population density declines rapidly (Fig. 3). Irrigation during and following nematode application increases the survival and efficacy of nematodes (Downing, 1994); however, large numbers of nematodes remain near the soil surface and die (Duncan and McCoy, 1996). Although recycling of exotic nematodes has been detected in experimental plots in the field, the level of long-term insect management does not appear to be significant. In a greenhouse experiment in which S. riobrave were applied at various intervals to potted citrus seedlings that were infested repeatedly with neonate larvae of D. abbreviatus, fibrous root weights of trees increased directly with the number of nematode applications (Duncan and McCoy, unpublished). Compared to trees not infested by weevils, fibrous roots of infested trees were reduced significantly even when treated monthly with nematodes. These data suggest that very limited feeding by the insect is likely to reduce fruit yield by diverting carbohydrates to fibrous root growth, and that low persistence by the nematode requires frequent application to mitigate the problem.
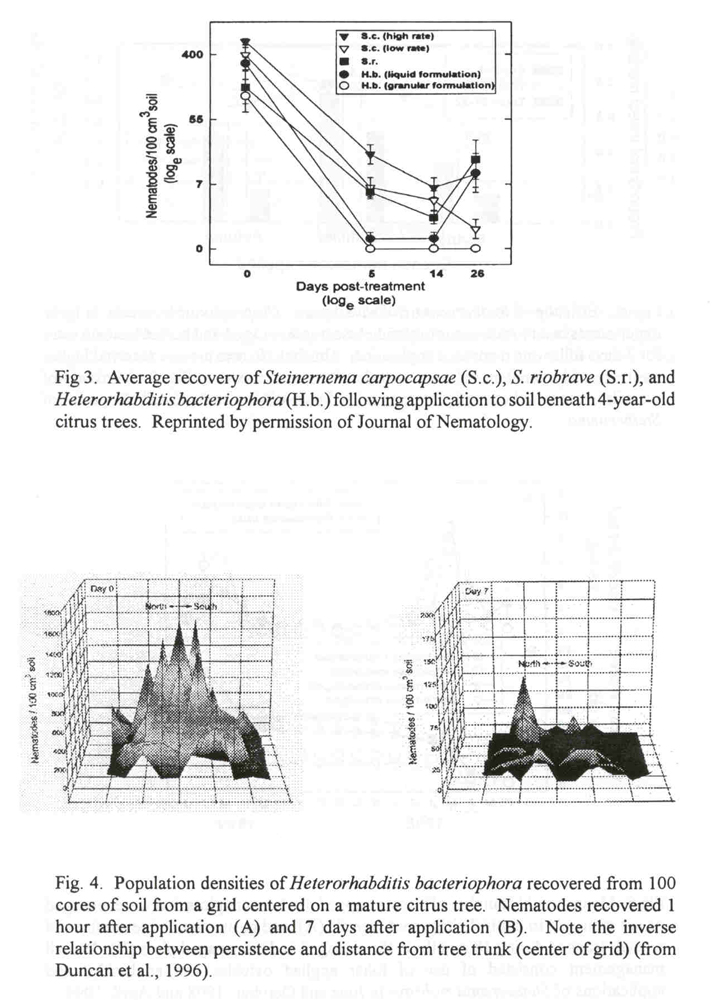
Time of day is also important when scheduling nematode applications (Fig. 4). Nematodes applied beneath the canopy of a tree survive in direct proportion to their proximity to the tree trunk where evaporation of soil moisture and exposure to ultraviolet radiation are least (Duncan et al., 1996; Molyneaux and Bedding, 1984; Gaugler and Boush, 1978). Application of nematodes in the evening provides the longest possible time for their establishment before being exposed to desiccation and sunlight.
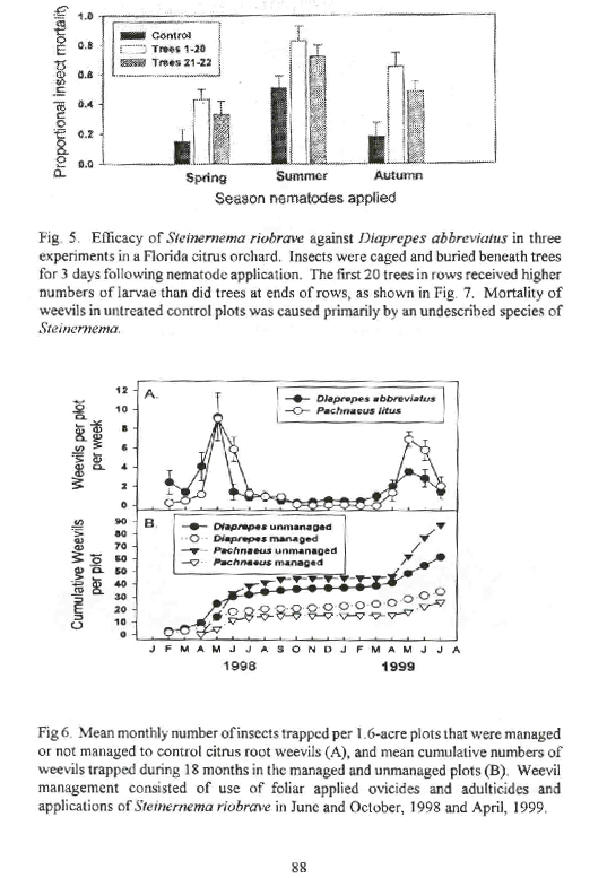
Indigenous entomopathogenic nematodes are generally found to have highest activity during summer months in Florida (Beavers et al., 1983) and elsewhere (Doucet and Giayetto, 1998). These natural control agents appear to be key mortality factors regulating the population dynamics of citrus root weevils. Recent experiments in Florida have found natural, nematode-induced mortality of D. abbreviatus during summer to be as high as 40-50% after just 3 days in the soil (Fig. 5). Increased activity of indigenous nematodes coincides with the onset of the characteristic seasonal depression in numbers of insects emerging from soil (Fig. 6). Important questions regarding these as yet undescribed species include understanding their incidence throughout the industry, whether the level of natural control is dependent on root weevil density, and whether they are good candidates for augmentation by virtue of their ability to persist under Florida conditions, or for other reasons.
Insect Population Biology
Because nematodes attack insects in the soil and show little evidence of significant persistence, it is generally assumed that nematodes will have the greatest effect if applied when numbers of insects in soil are highest. Emergence from soil of D. abbreviatus and another species of root weevil, Pachnaeus litus, is seasonal in Florida, with maximum emergence in late spring (Fig. 6A). The data in Fig. 6A are from Tedders traps which are placed under the tree canopy and require insects to climb a dark colored base into an elevated trap. Tedders traps are more efficient than cone shaped traps placed on soil beneath tree canopies, and seasonal patterns of adult activity are the same for both types of trap (McCoy, unpublished). There are no comparable census data for densities of insect larvae in soil; however, the adult census data suggest that the rate of egg deposition in the tree canopy increases in early summer. Thus, by autumn the surviving larvae in the soil have likely reached a maximum density, because low winter temperatures greatly reduce ovipositional activity. As temperatures increase in the spring, larval and pupal development continues until adults emerge from the soil. Because larval development requires a minimum of 4 months, it is likely that most larvae which enter soil during mid-to-late summer emerge the following spring.
Growers generally do not apply more than 2 applications of nematodes per year for economic rather than empirical reasons. Based on the pattern of adult emergence from soil, there is general consensus that an application of nematodes in the autumn presents the parasites with their greatest opportunity to locate insect prey. A second application of nematodes in spring when soil temperatures are high enough for nematode activity, but before adult emergence occurs, is practiced by many growers and researchers (e.g. Bullock et al., 1999). To reduce the deposition of larvae into soil following a springtime nematode treatment, management of aboveground stages of the insect is recommended at peak adult emergence. It has also been suggested that an application of nematodes in summer, when rainfall and soil temperatures are highest, provides the nematodes with ideal conditions for parasitism at a time when natural control is highest (Knapp, 1998).
Clearly, more realistic management models are needed to determine the optimum number and timing of nematode applications. Such models will require a great deal of additional basic information on insect population dynamics, and on questions such as effect of insect developmental stage on rate of nematode infection, and on positive or negative interactions between indigenous and exogenous nematode species.
Application Methods
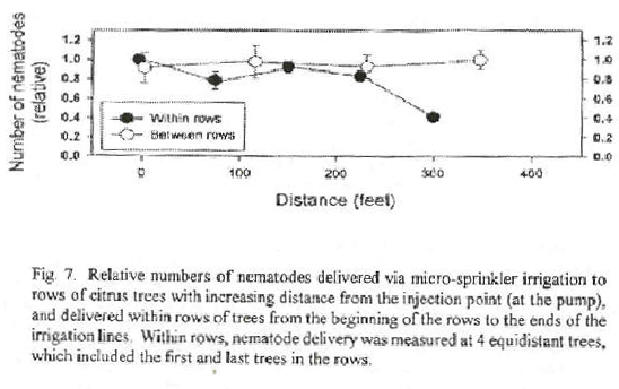
Entomopathogenic nematodes are applied to citrus either with herbicide application equipment or via under-tree, low-volume irrigation systems. The latter method has the advantages of eliminating the cost of driving equipment through the orchard and of depositing nematodes only in irrigated soil. However, the spatial pattern of nematode deposition throughout a grove is less consistent when applied by irrigation than with tractor-driven equipment. Nematodes tend to settle to the bottom of irrigation lines, particularly when the flow rate is low as in drip-irrigation systems (Conner et al., 1998). Micro-sprinkler irrigation systems have higher flow rates and deliver nematodes more uniformly. Numbers of nematodes delivered to tree rows is reasonably uniform with distance from the injection point (Fig. 7). A similar pattern is seen within the tree rows, except that trees at the very ends of rows receive significantly fewer nematodes, due to changes in water flow as water reaches the ends of lines. In preliminary experiments, efficacy of S. riobrave against D. abbreviatus was only measurably affected at the ends of tree rows (Fig. 5). This deficiency can be corrected by adding additional emitters or emitters with increased water delivery at the ends of rows.
Various common-sense factors should be considered with regard to application equipment. Holding/mixing tanks should be thoroughly cleaned of nematode-detrimental chemical residues from previous operations. Nematodes should never be combined with other chemicals to be applied to trees. The pH of delivery water should not be excessively low or high. Artesian well water lacks sufficient oxygen for nematodes and should not be used. Pumps that generate excessive heat should not be used for injecting nematodes into irrigation systems or for maintaining nematodes suspended in holding tanks.
Application Rate
Label rates for S. riobrave and H. bacteriophora in citrus are 200 million and 100 million nematodes per acre, respectively. However, the actual rate of application varies with tree age, because the under-canopy surface area of young trees is an order of magnitude less than that of mature trees. Thus, reported application rates (using label recommendations) vary from more than 200 to fewer than 20 nematodes per cm2 soil surface. At the higher rates, short-term efficacy against D. abbreviatus was found to be very high (85-95%; Duncan et al., 1996; Duncan and McCoy, 1996; Bullock et al., 1999). On mature trees at the lower application rates, efficacy has tended to be much lower (Figs. 5 and 6B). Results shown in Fig. 5 are typical of two ongoing experiments and indicate a dosage response to S. riobrave rate. Efficacy of the nematode against D. abbreviatus larvae buried beneath trees at the ends of rows was consistently lower than efficacy measured beneath other trees in those rows. Trees at the ends of rows were shown to receive far fewer nematodes than other trees (Fig. 7). These modest estimates of short-term efficacy are confirmed by the cumulative numbers of D. abbreviatus and Pachnaeus litus that were trapped in those plots during an 18 month period (Fig. 6B). Moreover, nematode applications in this experiment were used in combination with other tactics to manage the insect. It should be noted that these results are in marked contrast to those of a similar study in which cumulative numbers of adult D. abbreviatus trapped during one year were reduced by up to 95% by the application of the label rate of S. riobrave (Bullock et al., 1999). Nevertheless, trees in the study by Bullock et al. (1999) were relatively young (7 years), and nematodes were applied by sprinkling can, suggesting that the area treated was small.
The recommended application rate for S. riobrave has consistently produced very high short-term efficacy against root weevils in young trees. Results in mature trees, where the effective application rate is lower, have been consistently measurable, but variable in magnitude. The data suggest that further research to determine an expected dosage-response based on surface area treated is warranted. In Florida, such trials should be conducted on the sandy soils in the central part of the state, and on the heavier soils along the coasts.
Conclusions
The nature of the life cycle of Diaprepes abbreviatus presents a serious and complex management challenge because insects are continually recruited from soil to the tree canopy and from the canopy to soil. Short-term suppression of either the aboveground or belowground stages of the insect, independently of one another, is unlikely to provide adequate control. However, in the absence of host resistance, IPM relies increasingly on non-persistent, narrow-spectrum tactics to manage insects with the least environmental disruption. Entomopathogenic nematodes have been found to have outstanding potential for use as a component of root-weevil IPM. Nevertheless, breaking the insect recruitment cycle requires a great deal of additional knowledge in order to intervene with the correct tactics at the appropriate time.
Future research should evaluate the relative contribution to overall insect control of current tactics to intervene in the above ground (mainly insecticides) vs the below ground (insecticides or nematodes) stages of the insect life cycle. Profitability of current IPM programs should be evaluated to provide a baseline for future management innovations. Population models and economic thresholds relating population density to root damage and damage to yield are urgently needed and require improved methods to monitor these insects. Finally the diversity of entomopathogenic nematodes should be exploited by further characterizing the biology and biocontrol potential of known and yet to be discovered indigenous and exotic species.
Selected References
1. Beavers, J. B., C. W. McCoy, and D. T. Kaplan. 1983. Natural enemies of subterranean Diaprepes abbreviatus (Coleoptera: Cucurlionidae) larvae in Florida. Environmental Entomology 12:840-843.
2. Bullock, R. C., R. R. Pelosi, and E. E. Killer. 1999. Management of citrus root weevils (Coleoptera: Curculionidae) on Florida citrus with soil-applied entomopathogenic nematodes (Nematoda: Rhabditida). Florida Entomologist 82:1-7
3. Conner, J. M., R. McSorley, P. A. Stansly, and D. J. Pitts. 1998. Delivery of Steinernema riobrave through a drip irrigation system. Nematropica 28:95-100.
4. Doucet, M. M. A. and A. L. Giayetto. 1998. Distribution of Heterorhabditis bacteriophora and Steinernema rarum (Heterorhabditidae and Steinernematidae) in cultivated fields in Oliva, Cordoba, Argentina. Nematropica 28:129.
5. Downing, A. S., S. G. Erickson, and M. J. Kraus. 1991. Field evaluation of entomopathogenic nematodes against citrus root weevils (Coleoptera: Curculionidae) in Florida citrus. Florida Entomologist 74:584-586.
6. Downing, A. S. 1994. Effect of irrigation and spray volume on efficacy of entomopathogenic nematodes (Rhabditida: Heterorhabditidae) against white grubs (Coleoptera: Scarabaeidae). Journal of Economic Entomology 87:643-646.
7. Duncan, L. W. and C. W. McCoy. 1996. Vertical distribution in soil, persistence, and efficacy against citrus root weevil (Coleoptera: Curculionidae) of two species of entomogenous nematodes (Rhabditida: Steinernematidae; Heterorhabditidae). Environmental Entomology 25:174-178.
8. Duncan, L. W., C. W. McCoy, and A. C. Terranova. 1996. Estimating sample size and persistence of entomogenous nematodes in sandy soils and their efficacy against the larvae of Diaprepes abbreviatus in Florida. Journal of Nematology 28:56-67.
9. Gaugler, R. and G. M. Boush. 1978. Effects of ultraviolet radiation and sunlight on the entomogenous nematode, Neoaplectana carpocapsae. Journal of Invertebrate Pathology 32:291.
10. Graham, J. H. and J. A. Menge. 1999. Root Diseases. In: Citrus Health Management, eds. L. W. Timmer and L. W. Duncan, pp 126-135. APS Press: St. Paul.
11. Kaya, H. K., T. M. Burlando, and G. S. Thurston. 1993. Two entomopathogenic nematode species with different search strategies for insect suppression. Environmental Entomology 22:859-864.
12. Knapp, J. L. 1998. 1998 Florida Citrus Pest Management Guide. Cooperative Extension Service - IFAS: Gainesville.
13. McCoy, C. W. 1999. Arthropod pests of citrus roots. In: Citrus Health Management, eds. L. W. Timmer and L. W. Duncan, pp 149-156. APS Press: St. Paul.
14. Molyneux, A. S. and R. A. Bedding. 1984. Influence of soil texture and moisture on the infectivity of Heterorhabditis sp. Di and Steinernema glaseri for larvae of the sheep blowfly, Lucilia cuprina. Nematologica 30:358-365.
15. Schroeder, W. J. 1990. Suppression of Diaprepes abbreviatus (Coleoptera: Curculionidae) adult emergence with soil application of entomopathogenic nematode (Nematoda: Rhabditida). Florida Entomologist 73:680-683.
16. Schroeder, W. J. 1994. Comparison of two steinernematid species for control of the root weevil Diaprepes abbreviatus. Journal of Nematology 26:360-362.
17. Shapiro, D. I., J. R. Cate, J. Pena, A. Hunsberger, and C. W. McCoy. 1999. Effects of temperature and host age on suppression of Diaprepes abbreviatus (Coleoptera: Curculionidae) by entomopathogenic nematodes. Journal of Economic Entomology (in press).
| Nematode | Percent Reductionb |
Reference | |||||
|---|---|---|---|---|---|---|---|
| S. riobrave | 110 | Watering can (2) | March/ Sept. | Indian River County | Diaprepes Pachnaeus | 98% 95% |
Bullock et al. (1999) |
| S. riobrave | 110 | Watering can (2) | March | Indian River County | Diaprepes Pachnaeus | 82% 80% |
Bullock et al. (1999) |
| S. riobrave | 110 | Watering can (1) | March | Indian River County | Diaprepes Pachnaeus | 85% 65% |
Bullock et al. (1999) |
| S. riobrave | 110 | Injection L.V.c irrigation (1) |
March | Indian River County | Diaprepes Pachnaeus | No control | Bullock et al. (1999) |
| S. riobrave | N.D. (2 million/ tree) |
Injection L.V.c irrigation (1) |
March | Indian River County | Diaprepes Pachnaeus | 100% 90% |
Bullock et al. (1999) |
| S. riobrave | N.D. (2 million/ tree) |
Injection L.V.c irrigation (1) |
March | Indian River County | Diaprepes Pachnaeus | 51% 100% |
Bullock et al. (1999) |
| S. riobrave | N.D. (1.6 million/ tree) |
Herbicide applicator (1) |
March | Indian River County | Diaprepes Pachnaeus | 98% 48% |
Bullock et al. (1999) |
| S. riobrave | 120 | Watering can (1) | May | Lake County | Diaprepes | 90% | Duncan et al. (1996) |
| S. riobrave | 250 | Watering can (1) | Oct. | Polk County | Diaprepes | 77-90% | Duncan and McCoy (1996) |
| S. riobrave | N.D. (3-9/cm3) |
Watering can | N.D. | Greenhouse pot test |
Diaprepes | 77-86% | Schroeder (1994) |
| H. bacter- iophora |
175-250 | Watering can (1) | May | Lake County | Diaprepes | 55% | Duncan et al. (1996) |
| H. bacter- iophora |
250 | Watering can (1) | Oct. | Polk County | Diaprepes | No control | Duncan and McCoy (1996) |
| H. bacter- iophora (Otinem) |
N.D. (7.8 million/ tree) |
(7 appl., in 3 yr) Injection via L.V. irrigation (1) |
Nov. | Indian River County | Diaprepes | No control | Adair (1994) |
| H. bacter- iophora (Otinem) |
127 255 637 |
Injection via L.V. irrigation (1) | Early spring | Lake County | Diaprepes | 83% 78% 69% |
Downing et al. (1991) |
| H. bacter- iophora (Otinem) |
127 255 637 |
Injection via L.V. irrigation (1) | Early spring | Lake County | Diaprepes | 72% 47% 56% |
Downing et al. (1991) |
| H. bacter- iophora (Otinem) |
127 255 637 |
Injection via L.V. irrigation (1) | Early spring | Osceola County | Pachnaeus | 17% 53% 76% |
Downing et al. (1991) |
| H. bacter- iophora (Otinem) |
160 | N.D. | March | Lake County | Diaprepes | 58% | Schroeder (1990) |
| H. bacter- iophora (Fl. strain) |
160 | N.D. | March | Lake County | 26% | ||
| aN.D. = no data. bEfficacy defined as the percent reduction in larvae in soil or adults emerging from soil. cL.V.= Low volume. |
|||||||